
Гаджеты
-
 1-3 Декабря 2009 г состоится научно-практическая конференция, посвященная развитию отечественной наноиндустрии.
1-3 Декабря 2009 г состоится научно-практическая конференция, посвященная развитию отечественной наноиндустрии. 1–3 декабря 2009 г. в г...
-
 10 Безумных гаджетов для отправки сообщений
10 Безумных гаджетов для отправки сообщений Стилус-насадка для носа — гаджет для тех, кто постоянно мечтал иметь лишний палец на лице...
-
 10 Безумных периферийных устройств для компьютера
10 Безумных периферийных устройств для компьютера Titan Sphere — продукт скоро разорившейся компании SGRL, неудавшаяся попытка сообщить новое слово в сфере джойстиков...
-
 10 Безумных японских изобретений
10 Безумных японских изобретений Раструбы для глазных капель разрешают совершенно верно прицелиться в глаз, в то время, когда необходимо его чем-то зака...
-
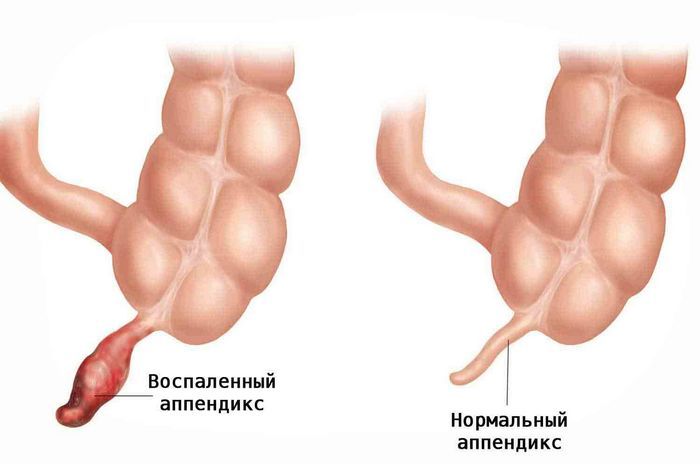 10 Бесполезных органов человека
10 Бесполезных органов человека Существуют ли в действительности ненужные органы? Вряд ли кому-то захочется расстаться со своим аппендиксом , пока он е...
-
 10 Важнейших стадий развития персональных компьютеров
10 Важнейших стадий развития персональных компьютеров «Мать всех демонов», 1968 год...
-
 10 Вариантов будущего через тысячу лет
10 Вариантов будущего через тысячу лет Будущее с инопланетянами — почему бы и нет? Кое-какие уверены, что инопланетяне уже среди нас...
Немного интересного:
-
 7 Настоящих людей-киборгов (фото и видео)
7 Настоящих людей-киборгов (фото и видео) Вместе с ноутбуками, планшетами и смартфонами мы взяли разработке, каковые снабжают множество усовершенствованных способ...
-
 Дружественный к велосипедистам городской грузовик
Дружественный к велосипедистам городской грузовик
Велосипед – самый чистый транспорт...
-
 Нттм-2011. подрастающая смена
Нттм-2011. подрастающая смена В № 9 отечественного издания был опубликован репортаж с выставки НТТМ-2011...
-
 Обнаружен новый объект во внешней части солнечной системы
Обнаружен новый объект во внешней части солнечной системы Спустя 10 лет с момента открытия Седны в самой дальней области отечественной звездной совокупности найден второй объект ...
-
 Полная мутация человеческого генома уже близко
Полная мутация человеческого генома уже близко Так как на данный момент доступно много образцов геномов человека для изучения, то современные ученые в полной мере смог...
-
Реплисомы оказались взаимно «прозрачны» при терминации репликации
22.10.2013
 Hi-tech
Hi-tech -
Обстоятельством высокой скорости терминации репликации у позвоночных была «прозрачность» прохождения двух реплисом с противоположных цепей ДНК приятель мимо приятеля. Установить это удалось молекулярным биологам из Гарвардской медицинской школы посредством созданной ими новой модельной совокупности. Изучение размещено в Nature.
Механизм терминации репликации, другими словами завершения синтеза ДНК, до сих пор изучен достаточно не хорошо, по крайней мере по сравнению с другими стадиями репликации. Это связано, в первую очередь, с методическими сложностями. Дело в том, что напрямую синхронизировать терминацию репликации в различных клетках весьма не легко.
А без синхронизации нереально заметить процесс на молекулярном уровне, — по крайней мере без применения одномолекулярных способов (каковые не хорошо подходят в этом случае).
Плазмида Col E1 под микроскопом. Изображение: J. Inselburg, PNAS, 1974
Создать модельную совокупность терминации репликации получалось до сих пор лишь у бактерий, дрожжей и у эукариатических вирусов (SV40).
") Но до сих пор было не ясно, как совершенно верно эти сведенья воспроизводят то, что происходит на протяжении работы собственной ДНК-репликазы высших эукариот.
Но до сих пор было не ясно, как совершенно верно эти сведенья воспроизводят то, что происходит на протяжении работы собственной ДНК-репликазы высших эукариот.Модельная совокупность, которую создали молекулярные биологи из Гарварда, является плазмидой , в которую встроен сайт посадки для 16 репрессоров из лактозного оперона (протяженность сайта образовывает 490 нуклеотидов). Эти репрессоры выступали в роли механических зажимов: они разрешали временно остановить продвижение реплисом из двух противоположных репликационных вилок.
После этого, при добавлении особого реагента (IPTG), репрессорные «зажимы» в один момент «спадали» с ДНК во всех клетках и репликация синхронно завершалась. Это разрешало отследить все стадии процесса терминации стандартными молекулярно-биологическими способами (радиоактивным электрофорезом в агарозе и иммунопреципитацией хроматина).
Взяв такую совокупность, авторам удалось продемонстрировать, что при столкновении репликационных вилок не происходит понижения скорости синтеза ДНК, как предполагали ранее кое-какие модели (обстоятельством этого назывались топологические затруднения). Движущиеся друг на друга реплисомы совсем не «ощущают» друг друга впредь до синтеза последнего нуклеотида, кроме того в момент расхождения цепей матричной ДНК.
Синтез завершается с присоединением лидирующей цепи одной репликационной вилки к отстающей цепи противоположной вилки. Затем реплисомы диссоциируют с ДНК. Но геликазы (белки, распутывающие ДНК перед движущейся полимеразой) двух репликационных вилок остаются связанными с нуклеиновой кислотой впредь до полного лигирования всех снова синтезированных распутывания и фрагментов (декатенации) двух дочерних молекул ДНК.
Последовательность различных стадий терминации репликации, установленная в новой работе. Изображение: James M. Dewar , Magda Budzowska, Johannes C. Walter, Nature, 2015
Наровне с трансляцией и транскрипцией репликация есть одним из центральных процессов, каковые исследует молекулярная биология начиная с 50-х годов прошлого века. Молекулярный механизм большинства из этих процессов установлен достаточно давно и количество аналогичных «белых пятен» в базисных процессах на сегодня весьма мало.
Случайные записи:
- Роснано выражает несогласие с рядом выводов, содержащихся в акте проверки компании счетной палатой
- Композит расширяет горизонты
Цитология. Лекция 55. Репликация ДНК
Похожие статьи, которые вам понравятся:
-
Ученые научились синтезировать молекулы и наноструктуры при помощи специальных искусственных белков
Ни для кого не есть секретом, что развитие некоторых областей науки и техники тесно связано с развитием нанотехнологий. И по сей день множество групп…
-
Ученые нашли новый метод управления светом лазера при помощи странностей некоторых законов физики
Несколько исследователей из Венского Технологического и Принстонского университета , найдя кое-какие странности в математических уравнениях,…
-
Новая система получения водорода из воды
Комплексы железа являются действенными катализаторами, содействующими фотохимического выделению водорода из воды. Водород есть многообещающим носителем…
-
Китайцы создали сверхпроводимость при помощи давления
Физики Поднебесной нашли необычное свойство у селенидов железа: при повышении давления вещество сперва теряют собственную сверхпроводимость, но после…